The General Plant Cell
We covered plant cells back in the chapter on Cell Processes, but we’ll provide a summary here.
- Plants have all the organelles animal cells have. Nucleus,
ribosomes, mitochondria, endoplasmic reticulum, Golgi apparatus, etc.
- Plants have chloroplasts. Chloroplasts are special organelles
that contain chlorophyll and allow plants to carry out photosynthesis.
- Plant cells can sometimes have big vacuoles for storage.
- Plants are surrounded by a rigid cell wall made of cellulose,
in addition to the cell membrane that surrounds animal cells. These
walls provide support.
Types of Plants
Just as there are many different kinds of animals, there are also many
different kinds of plants. The earliest plants had no special vascular
tissues devoted to transport, meaning they could not grow to great
heights because they couldn’t transport necessary liquids and minerals
over long distances. These plants are called nonvascular or
nontracheophyte. Tracheophytes, also called vascular
plants, do have special vascular tissues for transport.
Vascular Tissues
Vascular tissues are composed of specialized cells that create “tubes” through which materials can flow throughout the plant body. These vessels are continuous throughout the plant, allowing for the efficient and controlled distribution of water and nutrients. In addition to this transport function, vascular tissues also support the plant. The two types of vascular tissue are xylem and phloem.
Leaves
Leaves are the sites of photosynthesis in plants. The leaves’ broad,
flattened surfaces gather energy from sunlight, while apertures on
their undersides bring in carbon dioxide and release oxygen. On its
two exteriors, the leaf has layers of epidermal cells that secrete a
waxy, nearly impermeable cuticle to protect against water loss
and fungal or bacterial attack. The only way gases can diffuse in or
out of the leaf is though stomata, small openings on the
underside of the leaf. The opening or closing of the stomata occurs
through the swelling or relaxing of guard cells. If the plant
wants to limit the diffusion of gases and the transpiration of water,
the guard cells swell together and close the stomata.
The tissues between the epidermal cells are called mesophyll.
The mesophyll can be further broken down into two layers, the
palisade layer and the spongy layer. Both layers are packed
with chloroplasts, the factories of photosynthesis. In the palisade
layer, chloroplasts are lined in columns just below the epidermal
cells to facilitate the capture of light. In the spongy layer, cells
are less ordered and more diffuse, leaving large intracellular spaces
that facilitate the exchange of carbon dioxide and oxygen.
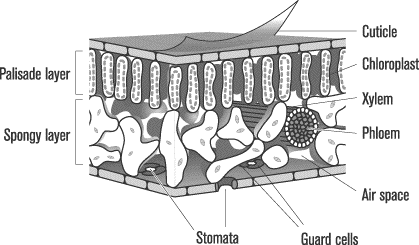
Overall, it is to the plant’s advantage to maximize the gas exchange and sunlight trapping surface area while keeping leaf thickness to a minimum so that once gases enter the leaf through the stomata, they can diffuse easily throughout the leaf cells.
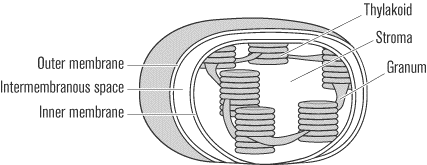
The Chloroplast
Like mitochondria, chloroplasts consist of a double membrane layer with
an intermembrane space between. The inner membrane is folded into
multiple stacks of flattened, disk-shaped compartments. Each such
compartment is called a thylakoid, and a stack of thylakoids is
called a granum. The thylakoid membrane separates the
chloroplast interior into two very different compartments: the
thylakoid space inside the thykaloids where the photosynthetic
chlorophyll resides and the stroma, a fluid that lies
outside the stacked disks and takes up the rest of the organelle.
Photosynthesis
In photosynthesis, plants (and other photosynthetic autotrophs) use
the energy from sunlight to create the carbohydrates necessary for
cell respiration. More specifically, plants take water and carbon
dioxide and transform them into glucose and oxygen:
6CO2 + 6H2O + light energy ? C6H12O6
+ 6O2
This general equation of photosynthesis represents the combined effects of two different stages. The first stage is called the light reaction, since it is dependent on light. The dark reaction, the second stage, does not need light.
The Light Reaction
The light reaction, also called the light-dependent reaction, takes place within the thylakoid spaces of the grana in the chloroplast. When sunlight strikes the chlorophyll contained within the thylakoid spaces, electrons become excited and infused with energy, and they are transferred down an electron transport chain similar to the one found in aerobic respiration. The energy in the electrons is used to set up a proton gradient across the membrane of the thylakoid spaces. Protons flowing back across the thylakoid membrane according to the concentration gradient are harnessed to produce ATP and NADPH (the reduced form of NADP). As a by-product of this process, molecules of water are split into molecular hydrogen and oxygen. The plant needs the hydrogen to produce ATP. The oxygen you’re breathing right now is some of that waste product.
The purpose of the light reaction is to make the usable energy necessary to run the dark reaction.
The Dark Reaction
The dark reaction is also referred to as the light-independent
reaction, the Calvin cycle, or carbon fixation. The reaction
takes place in the stroma of the chloroplast.
In the dark reaction, the carbon from carbon dioxide is added to the five-carbon sugar ribulose bisphosphate (RuBP) to produce a six-carbon compound. This six-carbon sugar is immediately split into two three-carbon molecules, which in a chain reaction using the ATP and NADPH from the light reaction are modified to form glyceraldehyde 3-phosphate. The glyceraldehyde 3-phosphate can be synthesized into carbohydrates such as glucose, and it can also be synthesized back into ribulose biphosphate. One of the glyceraldehyde 3-phosphate molecules is made into carbohydrates, while the other molecules remain in the Calvin cycle to serve as raw materials for the next round of production.
Roots
The roots of a plant draw water and minerals from the soil and pass them upward through xylem and phloem to the stem and leaves. Roots are also responsible for storing the plant’s organic nutrients, which are passed downward from the leaves through the phloem. Radiating from the roots is a system of root hairs, which vastly increase the absorptive surface area of the roots. Roots also anchor the plant in the soil.
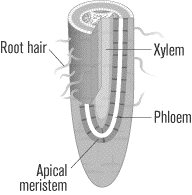
Growth in Vascular Plants
Vascular plants undergo two kinds of growth, primary growth and
secondary growth. Primary growth occurs in the apical meristems,
located at the tip of both root and shoot, and is mainly a growth of
vertical length. The meristems are regions of rapid mitotic
division, churning out cells like a factory. When a cell divides, one
of its offspring moves down into the plant body, where it elongates,
and the other remains in the meristem to divide again.
Secondary growth is a growth of thickness. Secondary growth is a
product of two different, though related, tissues, which both fall
under the umbrella-term lateral meristems. Vascular cambium
exists between xylem and phloem: on its inside the cambium produces
what is known as secondary xylem; on its outside it forms secondary
phloem. The primary xylem and phloem are pushed farther inward and
outward. The vascular cambium is more productive during the growing
seasons. During the rest of the year it creates little growth. This
phenomenon creates distinct rings of growth, each ring representing a
single growing season. By studying these rings, it is possible to
calculate the age of a plant, and it’s even possible to determine the
specific conditions of a particular growing season. The second lateral
meristem is called cork cambium and is responsible for the formation
of cork (bark), which replaces the epidermis to form the protective
covering of shoot and root.
Controlling Growth: Plant Hormones
Plant growth is controlled by plant hormones, which influence cell differentiation, elongation, and division. Some plant hormones also affect the timing of reproduction and germination.
- Auxins. The primary function of the auxin hormones is to
elongate plant cells in the stem. Auxins are also responsible for root
development, secondary growth in the vascular cambium, inhibition of
lateral branching, and fruit development.
- Kinins promote cell division and tissue growth in leaf, stem,
and root. Kinins are also involved in the development of chloroplasts,
fruits, and flowers. In addition, they have been shown to delay
senescence (aging), especially in leaves, which is one reason that
florists use cytokinins on freshly cut flowers.
- Gibberellins stimulate growth, especially elongation of the
stem, and can also end the dormancy period of seeds and buds by
encouraging germination. Additionally, gibberellins play a role in
root growth and differentiation.
- Ethylene controls the ripening of fruits. It also contributes
to the senescence of plants by promoting leaf loss and other changes.
Ethylene can bring buds and seeds out of dormancy, initiate flower
development, and promote radial (horizontal) growth in roots and
stems.
- Inhibitors restrain growth and maintain the period of dormancy
in seeds and buds.
Plant Behavior: Tropisms
When people think about plants growing, they generally think of them growing straight up or growing wider. But plants also display other types of growth in response to the stimuli within their environment. These responses to stimuli are called tropisms and are controlled by plant hormones. There are three main tropisms:
- hototropism is the tendency of a plant to move toward light.
Phototropism results from the rapid elongation of cells on the dark
side of the plant, which causes the plant to bend in the opposite
direction.
- Gravitropism refers to a plant’s tendency to grow toward or
against gravity. A plant that displays positive gravitropism will grow
downward, toward the earth. A plant that displays negative
gravitropism will grow upward, away from the earth. Most plants are
negatively gravitropic. Gravitropism is also controlled by auxin. In a
horizontal root or stem, auxin is concentrated in the lower half,
pulled by gravity. In a positively gravitropic plant, this auxin
concentration will inhibit cell growth on that lower side,
causing the stem to bend downward. In a negatively gravitropic plant,
this auxin concentration will inspire cell growth on that lower
side, causing the stem to bend upward.
- Thigmotropism, a reaction to touch, causes parts of the plant
to thicken or coil as they touch or are touched by environmental
entities. Tree trunks, for instance, grow thicker when exposed to
strong winds and vines tend to grow straight until they encounter a
substrate to wrap around.
Photoperiodism
Plants have a wide variety of flowering strategies involving what time of year they will flower and, consequently, reproduce. In many plants, flowering is dependent on the duration of day and night; this is called photoperiodism.
dependent on the duration of day and night; this is called photoperiodism.
All flowering plants have been placed in one of three categories with respect to photoperiodism: short-day plants, long-day plants, and day-neutral plants. Despite their names, however, scientists have discovered that it is the uninterrupted length of night rather than length of day that is the most important factor in determining when and whether plants will bloom. Short-day plants begin to bloom when the hours of darkness in a 24-hour period rise above a critical level, as when days shorten in the autumn. These plants include poinsettias, chrysanthemums, goldenrod, and asters. Long-day plants begin to flower when the duration of night decreases past a critical point, as when days lengthen in the spring and summer. Spinach, lettuce, and most grains are long-day plants. Finally, many plants are day neutral, which means that the onset of flowering is not controlled by photoperiod at all. These plants, which are independent both of night length and day length, include tomatoes, sunflowers, dandelions, rice, and corn.
Plant Reproduction
Plants can reproduce both asexually and sexually. Each type of reproduction has its benefits and disadvantages.
Asexual Reproduction
Through asexual reproduction, many plants can produce genetically
identical offshoots (clones) of themselves, which then develop into
independent plants. This process is also called vegetative
propagation. The many modes of vegetative propagation include the
production of specialized structures such as tubers, runners, and
bulbs. Grafting is an artificial form of vegetative propagation. The
advantages to this kind of asexual reproduction, which can occur
either naturally or artificially, stem from the fact that it can occur
more rapidly than seed propagation and can allow a genetically
superior plant to produce unlimited copies of itself without
variation.
Tubers
As seen in potatoes, tubers are fleshy underground storage structures composed of enlarged parts of the stem. A tuber functions in asexual propagation as a result of the tiny scale leaves equipped with buds that grow on its surface. Each of these buds can form a new plant that is genetically identical to the parent.
Runners
Runners are slender, horizontal stems that spread outward from the main plant, such as those found on strawberry plants. Entirely new plants can develop from nodes located at intervals on the runners; each node can give rise to new roots and shoots.
Bulbs
Bulbs such as onions and tulips are roughly spherical underground buds with fleshy leaves extending from their short stems. Each bulb contains several other buds that can give rise to new plants.
Grafting
In grafting, two young plants are joined, first by artificial means and then by tissue regeneration. Typically, a twig or bud is cut from one plant and joined to a rooted plant of a related species or variety. The twig or bud is called the scion, and the plant onto which is it grafted (and that provides the roots) is called the stock. The scion eventually develops into an entire shoot system. Grafting often allows horticulturalists to combine the best features of two different plants into one plant. Sometimes the stock and scion retain independent characteristics, and sometimes the stock alters the characteristics of the scion in some desirable way.
Sexual Reproduction in Plants
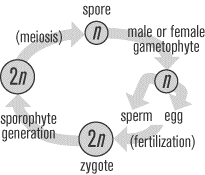
All plants undergo a life cycle that takes them through both haploid
and diploid generations. The multicellular diploid plant structure is
called the sporophyte, which produces spores through meiotic
division. The multicellular haploid plant structure is called the
gametophyte, which is formed from the spore and gives rise to the
haploid gametes. The fluctuation between these diploid and haploid
stages that occurs in plants is called the alternation of
generations. The way in which the alternation of generations
occurs in plants depends on the type of plant. In nonvascular plants,
the dominant generation is haploid, so that the gametophyte
constitutes what we think of as the main plant. The opposite is true
for tracheophytes, in which the diploid generation is dominant and the
sporophyte constitutes the main plant. The SAT II Biology only deals
with the specifics of the tracheophyte alternation of generations,
though nonvascular plants have a similar life cycle.
Next to display next topic in the chapter.
Practice Questions
Test Prep Lessons With Video Lessons and Explained MCQ
Large number of solved practice MCQ with explanations. Video Lessons and 10 Fully explained Grand/Full Tests.